LA
REGULATION DU CYCLE CELLULAIRE
| Accueil | B.Mol | Gen | T.P | Web |
| LA
REGULATION DU CYCLE CELLULAIRE |
Marie-Claude
Lebart, Jean Mariani
Gilles
Furelaud
Chapitre
8 - Le passage de G1 à la suite du cycle |
||
| sommaire : |
Au début de G1, la cellule contient le complexe Cycline D / Cdk 4 qui, une fois activé initie la phosphorylation de la pRb. Phosphorylée, la pRb libère E2F qui sert alors de facteur de transcription du gène de la cycline E. La fonction essentielle de la Cycline D / Cdk4 est donc d’activer, au début de la phase G1 (G1 précoce), la transcription du gène de la cycline E intervenant dans la phase suivante du cycle.
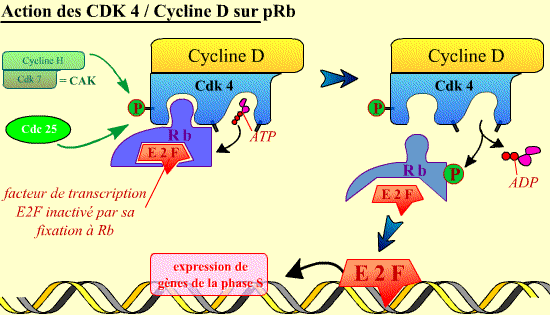
Mode d'action du complexe Cycline D / Cdk4 sur la protéine
Rb. Une fois activé, le complexe Cycline D / Cdk4 peut phosphoryler la
protéine Rb. Ceci permet alors la libération par cette
dernière du facteur de transcription E2F. Une fois libéré,
E2F se fixe sur l'ADN, activant l'expression de gènes codant pour des
protéines nécessaires au déroulement de la phase S. [Cliquer
sur l'image pour accéder à une explication
animée]
Une boucle de rétroactivation prépare le passage du point de restriction
Après la synthèse de la Cycline E, le complexe Cycline E / Cdk 2 pourra alors se former. Or, ce nouveau complexe a lui aussi pour protéine cible la p Rb qu’il phosphoryle. Il commande donc, par boucle de rétro-activation, l’amplification de la synthèse de sa propre cycline. Lorsqu’elle est hyperphosphorylée, la pRb libère le facteur E2F qui stimule alors la transcription du gène de la cycline suivante, la cycline A.
En réalité, il existe plusieurs versions de p Rb : p Rb, p 130, et p 107 qui séquestrent différents facteurs de transcription E2F (E2F1, 2, 3, 4, 5)
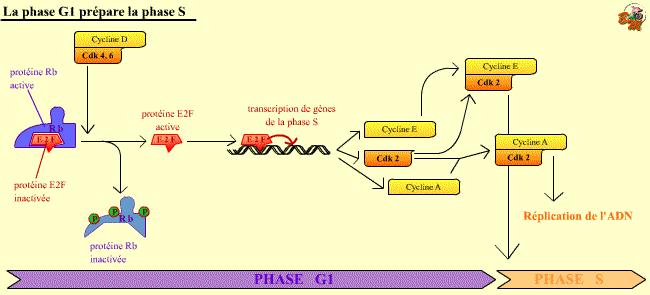
La phase G1 prépare la phase S. L'activation du
facteur de transcription E2F, grâce à la phosphorylation de la
protéine Rb, permet la synthèse de gènes de la phase S.
La transcription de Cdk 2 et de Cycline E, en particulier, permet d'augmenter
cette activation de E2F, ce complexe phosphorylant Rb . Ce mécanisme
permet à la cellule de préparer son entrée en phase S.
Le point de restriction, limite entre G1 précoce et G1 tardif
Le point de restriction désigne un moment spécifique de G1, avant
lequel les cellules en culture ont besoin de facteurs de croissance pour avancer
dans le cycle, et après lequel elles peuvent proliférer indépendamment
des stimuli mitogéniques.
Ce point a été mis en évidence en 1968 par A. Pardee.
Actuellement le point de restriction est utilisé pour subdiviser G1
en une phase G1 précoce et une phase G1 tardive.
Dans des cellules en culture, le point de restriction se situe 3 à 4
heures après la mitose. En revanche, la phase G1 tardive (du point de
restriction à la phase S) a une durée plus variable, de 1 heure
à 10 heures. Cette variabilité explique les différences
qui existent dans la longueur du cycle cellulaire.
On peut considérer que la cellule passe le point de restriction au moment où elle a, sous l’influence de facteurs de croissance, suffisamment synthétisé de cycline D et suffisamment mis en place de Cycline E / Cdk2 phosphorylant suffisamment de pRb. (La situation est probablement beaucoup plus complexe, avec intervention de signaux externes capables de réguler le nombre de cellules dans un organe, la taille d’un organe, ou même des signaux internes capables de mesurer le stade de croissance atteint par la cellule ...)
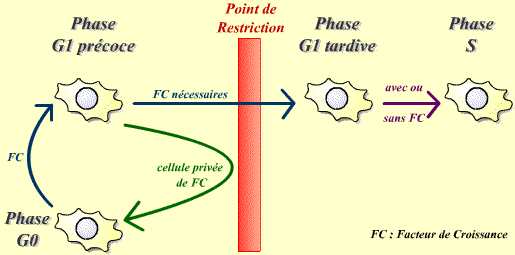
Le passage du point de restriction nécessite la présence
de facteurs de croissance.
Relation entre point de restriction et processus de surveillance G1 / S
Passé le point de restriction, la cellule commence irréversiblement un cycle, sauf si elle détecte une anomalie au niveau de l’ADN ; dans ce cas, le processus de surveillance G1/S est enclenché. Ainsi le processus de surveillance peut retenir la cellule en phase G1, c’est ce qu’illustre l’animation suivante.
Relation entre point de restriction et
processus de surveillance G1 / S. D'autres voies interviennent,
en plus de la p53 : voir le passage G1 / S. |
Protéines |
Partenaires ou cibles |
Rôles clés |
| Cdk | ||
| Cdk 4 | Cycline D | Fonctionne pendant G1 pour phosphoryler pRb. Directement induite par c-Myc |
| Cdk 6 | Cycline D | Phosphoryle pRb |
| Cdk 2 | Cyclines E et A | Fonctionne de G1 tardif à G2. Phosphoryle pRb et d'autres substrats comme les histones. Impliquée dans la duplication du centrosome et la réplication de l'ADN quand elle est liée avec la Cycline A |
| Cyclines | ||
| Cycline D | Cdk 4 et Cdk 6 | Quantité augmente en G1 et reste importante pendant la suite du cycle cellulaire |
| Cycline E | Cdk 2 | Induite après la phosphorylation de pRb par Cdk 4 et 6 |
| Inhibiteurs de Cdk | ||
| p16 | Cdk 4 et Cdk 6 | Induite pendant la sénescence et dans des stress oncogéniques. |
| p21 | Toutes les Cdk | P53 dépendante, induite durant la sénescence. Son niveau fluctue pendant le cycle, avec un pic en G1. Inhibe Cdk 2. Son absence prédispose aux tumeurs chez la souris |
| pRB - E2F | ||
| pRb, p130, P107 | E2F et beaucoup d'autres | Contrôlent la transcription de protéines liées au cycle. Sont régulées par les Cdk 4, 6 et 2 |
| E2F | pRb, p130, p107 | Cibles = gènes des Cyclines E et A, de pRb, de Cdk 1, de l'ADN polymérase |
| Phosphorylations | ||
| CAK (Cdk 7/Cycline H) |
Toutes les Cdk sauf Cdk7 |
Régule l'activité de presque tous les complexes Cdk / Cycline |
| Wee 1 | Complexes Cycline / Cdk | Phosphoryle et inactive Cdk 1 et 2. Régulé par Cdk 1et polo K |
| Cdc 25 | Complexes Cycline / Cdk | Active les Cdk 4, 6, 2 et 1 en enlevant les phosphorylations inhibtrices aux stades S et G2/M. Régulée par Cdk et polo K |
| Protéolyse | ||
| SPC | Impliqué dans la protéolyse médiée par l'ubiquitine des Cyclines D et E, ainsi que de p16 et E2F | |
| |
© Copyright "Biologie et Multimédia" | |
Mise
en ligne : juin 2004 |
 |