| Accueil | B.Mol | Gen | Web |


La gastrulation est caractérisée par :-
- 1 - L'apparition des mouvements cellulaires coordonnés
(mouvements morphogénétiques).
- 2 - La ségrégation des trois tissus fondamentaux : l'ectoderme, à l'origine de l'épiderme, le mésoderme à l'origine des mésenchymes primaire et secondaire, et l'endoderme à l'origine du tube digestif.
- 3 - L'apparition d'une cavité secondaire digestive, l'archentéron.
- 2 - La ségrégation des trois tissus fondamentaux : l'ectoderme, à l'origine de l'épiderme, le mésoderme à l'origine des mésenchymes primaire et secondaire, et l'endoderme à l'origine du tube digestif.
- 1 - L'apparition des mouvements cellulaires coordonnés
(mouvements morphogénétiques).
Parallèlement, le rythme mitotique diminue et le cycle cellulaire s'allonge
-
- Analyse des mouvements cellulaires au cours de la gastrulation
-
- On peut classer les mouvements cellulaires mis en oeuvre au cours de la gastrulation de l'oursin en 4 catégories. Chacun de ces mouvements est lié à une étape de la gastrulation, soit, dans l'ordre chronologique :
a - Ingression du mésenchyme primaire.
b - Migration du mésenchyme primaire.
c - Inflexion du pôle végétatif dans le blastocèle, qui initie la formation de l'archentéron.
d - Elongation de l'archentéron et individualisation du mésenchyme secondaire.a - Ingression du mésenchyme primaire
Les prémisses de la gastrulation s'observent au niveau du pôle végétatif. A cet emplacement, l'épithélium s'applatit (Fig.29). Les cellules du pôle végétatif issues des micromères montrent des mouvements pulsatiles sur leur face interne qui aboutissent à la formation de filopodes.
-
- Simultanément, alors que l'ensemble des cellules de la blastula possède une forte affinité pour la couche hyaline externe héritée des granules corticaux, les cellules des micromères perdent cette affinité et renforcent de 100 fois leur affinité pour la lame basale et la matrice extracellulaire interne. La lame basale se rompt et les cellules passent de l'état épithélial à l'état mésenchymateux. Elles se dirigent dans la cavité du blastocoele au contact de la matrice extracellulaire pour former les cellules du mésenchyme primaire (Fig.30, 31 et 32).

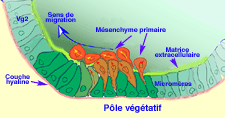
 Figure 30. Ingression des cellules du mésenchyme primaire d'oursin. Cliquez sur l'image pour l'agrandir.
Figure 30. Ingression des cellules du mésenchyme primaire d'oursin. Cliquez sur l'image pour l'agrandir.Figure 31. Schéma d'interpretation de l'ingression et de la migration des cellules du mésenchyme primaire d'après la figure 30. Cliquez sur l'image pour l'agrandir.
Figure 32. Schéma, avec les territoires présomptifs, de la jeune gastrula d'oursin au moment de l'ingression des cellules du mésenchyme primaire. Cliquez sur l'image pour agrandir.
- L'ampleur de cette migration reste relativement réduite dans la mesure où les cellules du mésenchyme primaire restent confinées latéralement dans l'hémisphère végétatif (Fig.33 et 34). Elles s'organisent en syncytium (fusion entre les cellules) dans lequel se dépose du carbonate de calcium à l'origine des spicules calcaires qui constituent le squelette larvaire (voir Fig.42).
-
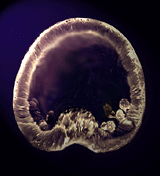
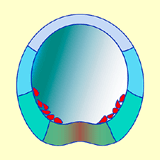 Figure 33. Jeune gastrula d'oursin avec les cellules du mésenchyme primaire en position latérale.
Figure 33. Jeune gastrula d'oursin avec les cellules du mésenchyme primaire en position latérale.Figure 34. Schéma, avec les territoires présomptifs, de la gastrula, au sortir de la migration des cellules du mésenchyme primaire. Cliquez sur l'image pour agrandir. -
Parallèlement à leur activité migratoire, les cellules du mésenchyme primaire développent de longues expansions cytoplasmiques ou filopodes qui ne paraissent pas être impliqués dans les mouvements de migration. Ces filopodes explorent la paroi du blastocoele et pouraient jouer un rôle dans la reconnaissance de la polarisation dorsoventrale de l'ectoderme. L'acquisition de ces informations permettrait d'établir une régionalisation au sein du mésenchyme primaire et aboutirait à l'élaboration des spicules paires qui marquent la symétrie bilatérale originelle des oursins (Fig.42).
-
-
- Une fois les cellules du mésenchyme primaire dans le blastocoele, le pôle végétatif est maintenant formé par les cellules du territoire Vg2 à l'origine du tube digestif. Celles-ci restent sous forme d'un épithélium cohérent de cellules adhésives. Leur pôle apical renferme des vésicules de sécrétion contenant un protéoglycane, la chondroitine sulfate, et est hérissé de microvillosités qui traversent la couche hyaline. Cette dernière, née du contenu des granules corticaux, sert d'ancrage aux cellules qui changent de forme. Par exemple, si on bloque les sites d'adhésion de ces cellules à la couche hyaline par des anticorps qui reconnaissent les protéines constitutives de la couche hyaline, les cellules s'arrondissent et le processus d'inflexion du pôle végétatif est inhibé.
- Un examen plus précis fait ressortir que la couche hyaline
est composée de deux sous-couches étroitement accolées,
la hyaline interne et la hyaline externe, impliquées dans les
mouvements d'inflexion du pôle végétatif que l'on
conçoit de la manière suivante.
Dans un premier temps, les vésicules de chondroitine sulfate déversent leur contenu dans la hyaline interne. Or, la chondroitine sulfate étant très hydrophile, la hyaline interne se gonfle d'eau et se dilate, inversement à la hyaline externe dont le volume ne change pas. La résultante de ces changements physiques conduit à des forces de tension entre les deux couches de hyaline. L'expansion latérale de la hyaline interne provoque une courbure concave vers le blastocoele et repousse les cellules du territoire Vg2, du pôle végétatif vers l'intérieur de l'embryon (Fig.35, 36 et 37).
Ce mécanisme peut être comparé à la courbure générée au sein d'une plaque formée de deux matières solidaires et possédant chacune un coefficeint de dilatation différent (pour plus d'explications voyez Fig. 36. Une animation suit la figure légendée). -


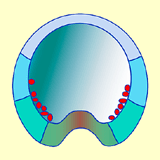 Figure 35. Mouvement d'inflexion du territoire Vg2 au pôle végétatif.
Figure 35. Mouvement d'inflexion du territoire Vg2 au pôle végétatif.Cliquez sur l'image pour l'agrandir.
Figure 36. Modèle pour expliquer la génération d'une courbure au sein de deux couches solidaires ayant chacune un coefficient de dilatation différent.Cliquez sur l'image pour l'agrandir.Figure 37. Schéma montrant les mouvements d'inflexion du pôle végétatif qui conduisent le territoire Vg2 à ébaucher la cavité de l'archentéron. Cliquez sur l'image pour agrandir.

-
b - Migration du mésenchyme
primaire
Les cellules en migration entrent en contact avec la matrice extracellulaire, sur la paroi du blastocoele. Plusieurs molécules jouent un rôle important dans ces événements. Deux d'entr' elles sont particulièrement importantes pour l'adhésion et la migration des cellules du mésenchyme primaire, la fibronectine, protéine de 220kDa, constituant majeur de la matrice extracellulaire, et des protéoglycanes sulfatés associés à la surface des cellules du mésenchyme primaire en migration. Il a été montré que pendant la gastrulation, l'affinité des micromères pour la fibronectine s'accroît considérablement.
Cet évènement est dèjà perceptible chez l'oursin sur la figure 33 et particulièrement bien visible dans l'embryon d'étoile de mer (Fig.38).
-
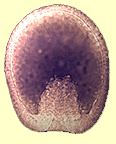
Figure 38. Gastrula d'étoile de mer dont l'inflexion du pôle végétatif est en cours. Cliquez sur l'image pour agrandir. -
- L'élongation de l'archentéron est assurée par l'aplatissement des cellules qui composent ses parois. De plus, des mouvements d'intercalation cellulaire réduisent le diamètre de l'archentéron pour en faire un tube étroit et allongé. On parle de mouvements de convergence extention particulièrement nets chez l'étoile de mer (Fig.39).
-
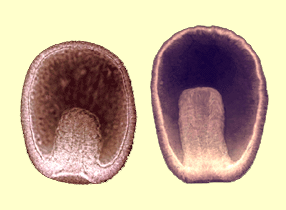
Figure 39. Deux stades d'élongation de l'archentéron chez l'étoile de mer.
Un autre mécanisme peut contribuer à cette élongation. Les cellules situées à l'apex de l'archentéron émettent des filopodes, parfois très longs, puisqu'ils vont à la rencontre du toit du balstocoele (Fig.40).
-

Figure 40. Mise en évidence des cellules du mésenchyme secondaire. Cliquez sur l'image pour agrandir.
Une fois cette connexion assurée, les fipodes se contractent, exerçant une force de traction qui contribue à l'allongement de l'endoderme vers le pôle animal. Ces mouvements concernent toujours le territoire Vg2 (Fig. 41).
-
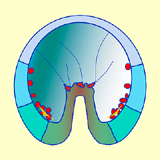
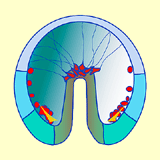
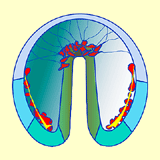 Figure 41. Elongation de l'archentéron et formation du mésenchyme secondaire. Cliquez sur les images pour agrandir.
Figure 41. Elongation de l'archentéron et formation du mésenchyme secondaire. Cliquez sur les images pour agrandir. - Il est possible de mettre en évidence expérimentalement le rôle des longs filopodes émis par les cellules du mésenchyme secondaire. La rupture des filopodes à l'aide d'un faisceau laser stoppe la phase terminale de l'élongation de l'archentéron. Dans ces conditions, son allongement ne dépasse pas les 2/3 de sa longueur finale. Cependant si on laisse seulement quelques cellules du mésenchyme secondaire en place, l'allongement de l'archentéron s'effectue mais plus lentement que chez les embryons contrôles. Les étapes successives de la gastrulation montrant l'évolution des territoires présomptifs vous sont présentées dans une animation.
- Dans la suite du développement, alors que l'archentéron approche du pôle animal, les cellules du mésenchyme secondaire se séparent de l'endoderme et se dispersent dans le blastocele où elles constitueront des organes mésodermiques coelomiques tel que l'hydrocoele (voir le chapitre sur l'organogenèse larvaire).
-
- Au total, l'embryon parvenu au terme de la gastrulation possède une organisation concentrique où l'on retrouve l'endoderme en position profonde, c'est le tube digestif primitif ou archentéron. L'épiderme constitue le feuillet limitant externe de l'embryon. Quant au mésoderme, il est représenté par les cellules des mésenchymes primaire et secondaire situées dans les espaces laissés libres entre l'épiderme et l'archentéron. Latéralement, les spicules peuvent être mis en évidence au microscope à fond noir (Fig.42).
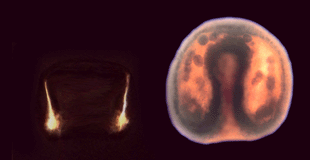 |
| Figure 42. Mise
en évidence des spicules au stade gastrula âgé.
Cliquez sur l'image pour l'agrandir. On peut mettre les deux structures
en coincidence grâce à une animation. |
![]()
