| Accueil | B.Mol | Gen | Web |


Chez les oursins, les sexes sont séparés. A la saison de la reproduction, les gamètes sont émis au niveau du pôle aboral. Hors de l'eau, les produits génitaux s'accumulent en formant une masse orange chez les femelles et blanche chez les mâles (Fig. 3).
|

|
|
Au laboratoire, les produits génitaux peuvent être émis spontanément, notamment, lors de la période de reproduction. C'est le cas sur la figure 3. Il est également possible de provoquer la ponte en injectant, à travers la région orale, de l'acetylcholine 0,1 mol.L-1, ou du KCl 0,5 mol.L-1. Il suffit alors de placer les oursins sur un récipient (par exemple, un Bécher ) rempli à ras bord d'eau de mer, le pôle aboral vers le bas de façon à ce qu'il soit immergé. On ne tarde pas, alors, à observer la lente descente des produits génitaux vers le fond du récipient où ils s'accumulent (Fig 4).
|

|
|
Cliquez sur l'image pour l'agrandir. (Photo Gérard Peaucellier). |
Les produits génitaux proviennent des gonades. Obéissant à la symétrie pentaradiée de ces animaux, les gonades sont au nombre de 5 et disposées de manière rayonnée par rapport à l'axe pôle oral-pôle aboral (Fig. 5).
|

|
|
|
![]()
-
- a) L'ovocyte
- L'ovocyte mesure un dizième de millimètre. Sa pauvreté en substances de réserve ou vitellus le qualifie d'oligolécithe.
- Dans l'ovaire, l'ovocyte possède un noyau de grande taille appelé vésicule germinative. Par réfringence, on discerne le nucléole (Fig.6 et 7).
-


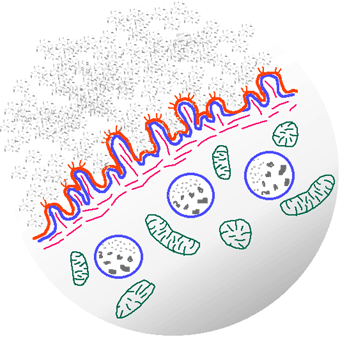
Figure 6. Ovocyte de Paracentrotus avant la maturation Figure 7. Schéma d'interprètation de la structure d'un ovocyte d'oursin L'ovocyte est limité par une membrane plasmique garnie de microvillosités et comportant des canaux ioniques. Sous cette membrame, le cortex cytoplasmique renferme de l'actine et des vésicules de type lysosomes. Ce sont les granules corticaux qui sont au nombre de 15000 environ. Chacun mesure 1 micron de diamètre. Leur contenu hétérogène examiné au microscope électronique, est dense aux électrons (Fig. 8).
-
-
Figure 8. Détail de la région corticale d'un ovocyte avant la fécondation. Cliquez sur la figure pour voir les légendes.
-
-
La surface externe de la membrane plasmique est recouverte de glycoprotéines assemblées en une membrane vitelline. Avant la fécondation, la membrane vitelline est étroitement accolée à la membrane plasmique. En plus de ces glycoprotéines, la membrane vitelline porte des protéoglycanes qui jouent le rôle de récepteurs d'une molécule qui sera présente à la surface du spermatozoïde au moment de la fécondation : la bindine.
La membrane vitelline est recouverte par les cellules folliculaires. Celles-ci sont elles-même recouvertes par une gangue comportant des glycoprotéines de haut poids moléculaire. L'une d'entr' elles riche en fucose- sulfate (FSG) joue le rôle de ligand pour des récepteurs présents sur la membrane plasmique du spermatozoïde au moment du contact de ce dernier avec la gangue.
Le cytoplasme de l'ovocyte comprend également des réserves informatives : les ARN ribosomiques, messagers et de transfert ; des molécules précurseurs des protéines et des acides nucléiques : acides aminés, nucléotides ; des protéines ; des lipides ; des réserves nutritives : le vitellus. En bref, l'ensemble du cytoplasme renferme toute la machinerie nécessaire aux opérations courantes de la physiologie cellulaire, la traduction des protéines, la transcription des ARN et surtout la réplication de l'ADN qui sera fortement sollicitée après la fécondation, lors de la période de clivage au rythme mitotique intense.Chez l'oursin Paracentrotus, l'ovocyte ovarien présente une pigmentation orangée uniforme. D'ailleurs, les oursins femelles à maturité se reconnaissent par l'émission de leurs produits génitaux orangés. Le pigment se rassemblera sous forme d'un anneau dans l'hémisphère végétatif après la fécondation. C'est le signe extérieur de la polarité axiale pôle animal-pôle végétatif. On parle alors d'hémisphère animal non pigmenté et d'hémisphère végétatif pigmenté (Fig.9).
-

Figure 9. L'ovocyte non fécondé de Paracentrotus possède une coloration orange clair uniforme (à gauche). Après la fécondation, le pigment orange se rassemble dans l'émisphère végétatif (à droite). Les globules polaires 1 et 2 ont été émis au moment de la maturation.
-
- La méiose a lieu au moment de l'ovulation lors de la rupture de l'enveloppe folliculaire. Les cellules folliculaires secrètent une hormone, la 1-méthyladénine, qui déclenche les divisions successives de la méiose. L'ovocyte émet alors consécutivement ses deux globules polaires au niveau du pôle animal, avant la fécondation. L'ovotide ainsi obtenu est fécondable.
Le nombre d'ovocytes pondus par une femelle peut être de l'ordre de la dizaine de millions et représente un volume de 10 à 20 millilitres.
-
- b) Le spermatozoïde
-
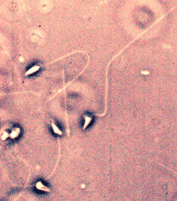
Figure 10. Suspension de spermatozoïdes de Paracentrotus. Cliquez sur la figure pour voir les légendes. - Le spermatozoïde est une cellule mobile, de petit taille (quelques microns), au cytoplasme très réduit (Fig .10). Il comprend trois parties principales :
- - la tête avec le noyau,
- - la pièce intermédiaire avec les centrioles et l'ensemble des mitochondries
- - la queue ou flagelle, organe de motricité.
- - la pièce intermédiaire avec les centrioles et l'ensemble des mitochondries
-
- Dans la tête, le noyau est formé de chromatine condensée liée à des protéines basiques, les histones et les protamines. L'enveloppe nucléaire dépourvue de pores est rigide et riche en lamine, une protéine de structure de la membrane nucléaire. La transcription est absente d'une telle structure aussi condensée.
- Devant le noyau, l'acrosome est formé par une vésicule de type lysosomial et contient des hydrolases ainsi que la bindine liée à la membrane de l'acrosome. Entre le noyau et l'acrosome, une réserve d'actine globulaire ou actine G (non polymérisée) servira à l'élongation du processus acrosomique (Fig. 11).
- Dans la tête, le noyau est formé de chromatine condensée liée à des protéines basiques, les histones et les protamines. L'enveloppe nucléaire dépourvue de pores est rigide et riche en lamine, une protéine de structure de la membrane nucléaire. La transcription est absente d'une telle structure aussi condensée.
-
Figure 11. Schéma du spermatozoïde d'oursin. Cliquez sur la figure pour voir les légendes. -
Figure 12. Schéma simplifié d'une coupe transversale dans un flagelle de spermatozoïde. Cliquez sur la figure pour voir les légendes.

|
| |
Le flagelle est formé d'axonèmes constitués majoritairement par des microtubules dont l'extrémité proximale est contiguë avec les centrioles de la pièce intermédiaire. Une coupe transversale du flagelle montre les axonèmes au nombre de 9 doublets disposés en cercle autour de 2 microtubules individuels centraux (Fig. 12).
|

|
| |
Les axonèmes sont formés par la polymérisation
des tubulines a et b, auxquelles s'adjoignent des protéines qui
possèdent un rôle dynamique telle que la dynéine.
Cette dernière, par son activité hydrolasique de l'ATP,
favorise le glissement des microtubules les uns par rapport aux autres
(phénomène de sliding), et par voie de conséquence,
l'animation du flagelle.
Autour des centrioles, une forte concentration de mitochondries est
là pour assurer l'alimentation énergétique de la
cellule.
Au total les produits génitaux, ovotides pour les femelles et
spermatozoïdes pour les mâles sont prêts à se
réunir pour effectuer la fécondation.
