
|


Pierre Clairambault (collaboration Gilles Furelaud)
1 - Introduction
De tous les réseaux de neurones qui constituent
l'encéphale des Vertébrés le système visuel
est l'un des mieux connus. On peut en établir l'évolution
anatomique en comparant entre-eux les plans de son organisation structurale
présentés dans chaque classe. Néanmoins, il faut
relativiser une telle démarche car, du fait de l'absence totale
de fossilisation du tissu nerveux, la dimension paléontologique
ne peut exister.
L'établissement d'une échelle de complexité des
structures visuelles chez les formes actuelles montre que ce sont les
Agnathes et les Amphibiens Urodèles qui présentent le plan
anatomique le plus simple (et que le plus élaboré est celui
des Primates). Entre ces deux extrêmes on peut observer certaines
ressemblances, par exemple entre les Téléostéens
et les Reptiles, ce que l'on peut interpréter comme le résultat
d'un phénomène de convergence évolutive en relation
avec une adaptation poussée de la fonction visuelle bien que dans
des milieux différents.
2 - La rétine
Embryologiquement, c'est une expansion du prosencéphale en
direction latérale ; il convient donc de l'interpréter
comme un appendice encéphalique et non pas comme un dérivé d'une
simple placode (épaississement) ectodermique comme c'est le cas
pour les autres organes sensoriels de la tête.
Dès le début de sa morphogenèse, l'oeil est composé
de deux feuillets cellulaires séparés par un ventricule
: la couche pigmentée et la couche nerveuse. Dans cette dernière
se différencient 5 types principaux de neurones répartis
en trois couches :
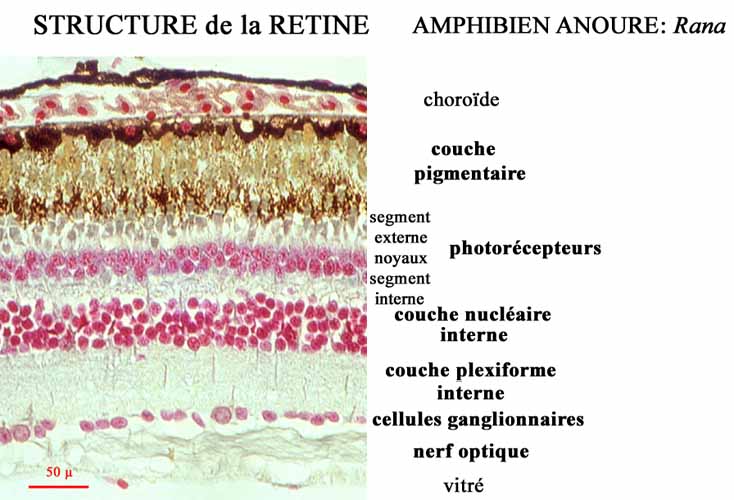 |
- La "granuleuse externe" (photorecepteurs dirigés vers la périphérie de l'oeil)
- La "granuleuse interne" (bipolaires, horizontaux et amacrines)
- La "couche des ganglionnaires" dont les axones (constituant le nerf optique) rejoignent les centres primaires.
- Ces couches, dites aussi nucléaires alternent avec
deux couches plexiformes,
externe et interne, composées de nombreuses synapses entre
les neurones cités.
Il faut aussi ajouter deux autres types cellulaires au rôle encore mal connu : les neurones interplexiformes et les neurones ganglionnaires déplacés (vers la "granuleuse interne").
 |
La glie (présente comme dans tous tissus nerveux) est représentée par des astrocytes (très communs dans le névraxe)
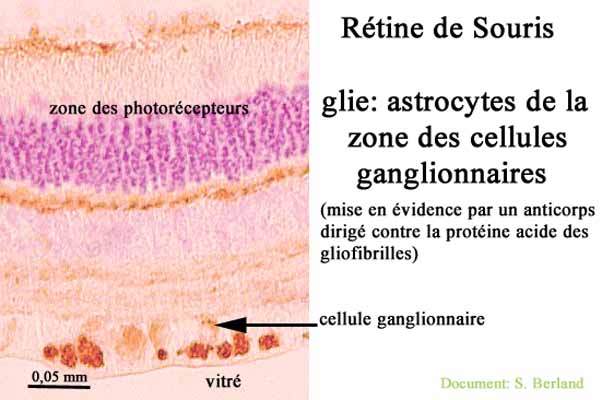 |
et les cellules de Müller spécifiques à la rétine.
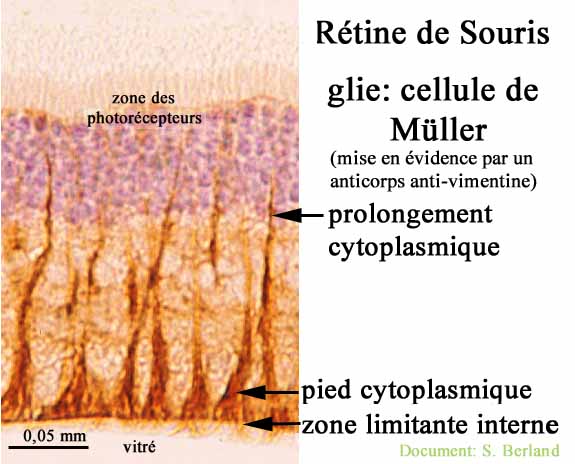 |
Cette extrême variété histologique implique une grande complexité de l'organisation synaptique qui fait de la rétine le premier véritable centre de décodage du message visuel.
3 - Organisation générale du système visuel primaire
Sortis de la rétine, les axones des neurones ganglionnaires, réunis dans chaque nerf optique (droit et gauche), arrivent à un carrefour (le chiasma) situé sur la face ventrale du diencéphale ; là, dans chaque nerf ils se séparent en deux groupes :
- L'un continue son trajet linéaire en direction de la moitié encéphalique opposée,
- L'autre tourne à angle droit et reste du même côté que
l'oeil correspondant.
Ces groupes vont respectivement constituer la voie croisée (ou controlatérale) et la voie directe (ou ipsilatérale) que vous présente l'animation ci-dessous :
Chacune de ces voies se scinde en plusieurs tractus conduisant aux différents centres visuels primaires :
- Les deux centres optiques , responsables de la formation des images : le toit optique (mésencéphale dorsal) et le thalamus dorsolatéral (diencéphale) composé lui même de plusieurs éléments dont "cops genouillé" (chez les Mammifères c'est le relai imposé sur le trajet rétino-cortical).
- Le centre hypothalamique impliqué dans l'élaboration des comportements visuo-dépéndants.
- le centre accessoire (mésencéphale ventral), uniquement controlatéral, qui intervient dans les réflexes visuo-moteurs.
Il résulte de cette organisation anatomique qu'à un
objet vu par les deux yeux au même instant, il peut correspondre quatre
images au niveau du thalamus comme au niveau du toit optique. Une
telle correspondance, cependant, n'existe pas chez tous les Vertébrés,
dans la mesure où la présence de la voie directe n'est
pas générale (par exemple, elle est absente chez certaines
espèces de Téléostéens et de Reptiles ).
Chez les Mammifères la plus grande partie des informations
visuelles remonte jusqu'au cortex occipital. Chez les autres
Tétrapodes, si le toit optique est le centre visuel majeur,
le télencéphale peut quand même recevoir ce type
d'informations via le thalamus.
Remarque : à tout l'ensemble de ces voies optiques rétinofuges,
s'ajoute une discrète voie rétinopète chez les
Agnathes et les Sauropsidés, impliquant, dans ces deux groupes, la
possibilité
d'une régulation du fonctionnement rétinien par l'encéphale
("feedback").
Il est facile de concevoir qu'une utilisation optimale de la vision est
essentielle
à la plupart des animaux, ceci implique une grande précision
dans la construction du cablage rétino-central ; cette modalité architecturale
est nommée rétinotopie : il s'agit d'une projection
rigoureusement organisée du réseau des cellules ganglionnaires
de la rétine vers les centres optiques primaires, de telle façon
qu'à toute cellule ganglionnaire correspond un locus synaptique
central précis. Chaque cellule ganglionnaire captant les informations
issues d'un très petit groupe de photorécepteurs, le résultat
de l'organisation rétinotopique est la construction d'une carte
topographique des photorécepteurs, point par point, sur les centres
encéphaliques ; et puisque les photorécepteurs
"voient" les différentes sources de stimuli (personnages,
objets, décor, etc..) c'est finalement une représentation
cartographique du champ visuel qui se dessine sur la structure encéphalique;
on peut alors dire que le cerveau "voit" (à son tour)
l'environnement de l'observateur. Ces photographies (en réalité un
film) vont
être utilisées par l'animal - observateur dans l'accomplissement
de nombreux comportements fondamentaux parmi lesquels celui de la capture
des proies, phase essentielle du comportement alimentaire (voir plus
loin).
4 – Le système visuel primaire des Amphibiens : exemple d’une organisation simple.
4 – 1. Malgré un rôle très important de la vision dans leurs comportements, même les Anoures présentent une organisation simple qui peut être considérée comme un plan anatomique fondamental du système visuel des Vertébrés. Les deux voies (croisée et directe) sont complètes bien que la voie controlatérale soit bien plus fournie en fibres rétinofuges et qu'elle soit la seule à en envoyer un contingent vers le centre accessoire. Tous les centres (thalamiques, prétectaux et tectaux) interviennent dans deux comportements essentiels : prise des proies et évitement des prédateurs (dossier " Spécificité des connexions rétino-tectales chez la grenouille. Première partie).
4 – 2. La connaissance précise de l’anatomie du système visuel des Anoures a permis aux neuroéthologistes d’étudier de manière approfondie le rôle de la rétinotopie dans différents comportements ( Spécificité des connexions rétino-tectales chez la grenouille. Deuxième partie).
L’organisation rétinotopique du toit optique d’une grenouille permet à l’animal d’apprécier (avec une bonne précision) les événements qui se déroulent dans son champ visuel, par exemple, lors de la recherche de proies (comportement alimentaire) ( Spécificité des connexions rétino-tectales chez la grenouille. Quatrième partie).
Cette organisation se met en place progressivement au cours de la métamorphose par une disparition sélective des projections inadéquates et une concentration topographique des projections adéquates : un remarquable modèle de spécificité neuronique.
4 – 3. Dans l’ensemble des Amphibiens, on observe d’intéressants exemples de réduction (hypomorphie) des structures visuelles centrales.
Chez les Urodèles comme les Salamandridés, toutes les structures sont présentes, mais comme leur rétine possède environ 10 fois moins de cellules ganglionnaires que celle des Anoures, les différents centres de projection sont plus discrets. La réduction (en particulier chez les formes aquatiques permanentes) affecte surtout la voie ipsilatérale.
Chez les Gymniophones (Apodes) les cellules ganglionnaires sont 10 fois moins nombreuses que chez les Urodèles, l’hypoplasie centrale est donc très importante ; aussi, chez Typhlonectes (vie fouisseuse) la voie ipsilatérale, très incomplète, ne dépasse pas le thalamus et la voie controlatérale atteint un centre tectal pour le moins ténu !
4 – 4. Sans faire allusion à une possible affinité phylétique avec les Amphibiens,les Dipneustes (3 espèces en tout !) ont également un système visuel primaire peu développé en particulier chez le Protoptère (africain) qui ne présente aucune trace de voie ipsilatérale (tout comme chez le cousin amazonien : Lepidosiren). Du côté controlatéral, l’ensemble est complet, bien que réduit avec, cependant, un doute sur l’existence d’un centre hypothalamique. Contrairement à ces deux espèces la troisième (australienne), Ceratodus, est pourvue d’un système complet des deux côtés. Une telle hétérogénéité à l’intérieur d’un même groupe est difficile à interpréter (voir le dossier Dipneustes à propos de la construction générale de l’encéphale) ; peut être est-elle simplement le résultat d’une adaptation à différentes niches écologiques éloignées les unes des autres et donc à différents modes de vie.
5 – Le système visuel primaire des Téléostéens : vers plus de complexité
L'ordre des Téléostéens contient un grand nombre d'espèces et montre ainsi une variabilité morphologique importante, notamment dans l'organisation de l'architecture encéphalique.Parmi toutes les espèces on peut quand même trouver une organisation anatomique de leur système visuel en forme de dénominateur commun en s'adressant à la truite.
5 -1. Comme pour les Amphibiens, la mise en évidence de l'organisation du système visuel se fait grâce à des techniques histologiques (anatomie microscopique) dont le principe repose sur un marquage sélectif des fibres rétinofuges. On agit expérimentalement sur les cellules ganglionnaires en utilisant 3 types différents de techniques :
- soit une destruction microchirurgicale des péricaryons suivie d'un traitement histologique qui met sélectivement en évidence les axones en dégénérescence wallérienne (méthode de Fink et Heimer) ;
- soit une microinjection, dans le vitré, d'une molécule (le plus souvent la proline) marquée par un atome radioactif (C14 ou H3), ensuite, la molécule est rapidement incorporée dans le métabolisme des cellules ganglionnaires et le marqueur circule le long de l'axone entraînépar le flux axonique jusqu'aux boutons synaptiques, puis les coupes histologiques sont traitées par autoradiographie et étudiées en microscopie "en fond noir";
- enfin on peut aussi utiliser d'autres molécules associées à un marqueur qui, incorporées dans les fibres rétinofuges, sont révélées par des techniques appropriées (réactions enzymatiques, immunofluorescence, etc..).
5 -2. L'anatomie générale du système visuel est caractérisée par :
- l'absence totale de structures ipsilatérales;
- la présence d'un nombre de centres de projection nettement plus important que chez les Amphibiens, donc une plus grande compléxité de l'organisation anatomique;
- un toit optique très développé.
La truite, animal chasseur, possède une rétine histologiquement
complexe et riche en cellules ganglionnaires. Dès leur entrée
dans le diencéphale les nombreuses fibres rétinofuges dessinent
un large éventail dans lequel on reconnaît 3
faisceaux qui aboutissent respectivement, l'un au tectum, l'autre
au thalamus dporsal et le dernier à
l'aire préoptique. Les fibres thalamiques
se projettent aussi dans une structure nommée improprement "corps
genouillé" par analogie avec les Amphibiens.
A l'entrée ventrale du tectum, la voie tectale se dirige vers
le prétectum puis vers les 3
couches visuelles tectales : superficielle, intermédiaire
(de loin la plus importante) et profonde. La signification de cette ségrégation
n'est pas connue.
5 -3. La mise en évidence sur coupes histologiques de la zone de projection de chaque quadrant rétinien :ventral , nasal, dorsal et temporal , a éte obtenue après destruction microchirurgicale d'un quadrant rétinien, injection de proline tritiée puis passage des coupes en autoradiographie. L'observation des préparations montrent que pour chaque quadrant, sa zone de projection intéresse toute l'épaisseur du tectum (donc les 3 couches visuelles) indiquant ainsi que la cartographie rétinotopique est construite en 3 dimensions. La carte tectale ainsi obtenue se rapproche nettement de celle des Amphibiens (bien que chez la truite on note l'existence d'une zone centrale qui semblerait, aux erreurs d'expériences près, complètement absente de projection).
Le corps genouillé présente aussi une rétinotopie pour 3 quadrants, seul le quadrant ventral ne se projette pas (ou très peu) sur ce territoire thalamique.
L'étude de la mise en place de la rétinotopie au cours du développement montre qu'il existe 2 catégories de fibres rétinofiges. Un premier groupe établit des projections qui resteront fixes sur le tectum pendant tout le développement : il s'agit de l'ensemble naso-ventral qui aboutit au tectum postéro-médial. Le second groupe, temporo-dorsal commence à se projeter sur le futur tectum central; les terminaisons axoniques sont alors entraînées en direction rostrale avec les nouvelles cellules tectales : ainsi les modalités mécaniques de la croissance tectale influent précocement sur l'organisation de la rétinotopie.5 -4. Remarques sur le système visuel primaire des Actinoptérygiens
Bien que l'organisation du système visuel de la truite soit représentative de beaucoup d'espèces de Téléostéens, il faut quand même signaler que des fibres ipsilatérales (à destination du thalamus st du prétectum sont présentes chez le poisson-chat (Ictalurus ponctatus) et le hareng (Clupea harangus). Chez les formes d'Actinoptérygiens primitifs comme l'esturgeon (Acipenser) ou le lepisosté (Lepisosteus) la voie ipsilatérale est la règle; on y note aussi une projection massive dans le prétectum et l'absence d'un corps genouillé.
![]()
