Gravitropisme : mécanisme
Le rôle de la coiffe et des stattolithes
 |
 |
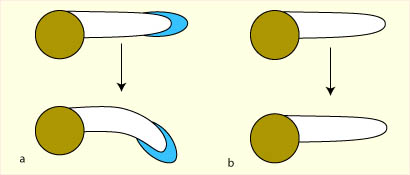
L'ablation de la coiffe d'une
racine couchée supprime sa réaction gravitropique. |
Après ablation de la moitié d'une coiffe, la croissance est inhibée du côté de la demi-coiffe restante. |
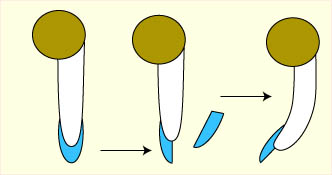
Dans l'apex de la racine, les cellules de la zone centrale de la coiffe (la columelle) sont appelées des "statocystes" car elles contiennent des amyloplastes appelés "statolithes" dont la position intracellulaire dépend de la position de la racine. De part leur forte densité par rapport aux autres organites cellulaires, ils se placent toujours vers le bas. Quand la racine est placée horizontalement, ils roulent pour se placer sur la face latérale de leur cellule.
Apex de racine. La région de la columelle est colorée. Schémas successisf du passage de la position verticale à la position horizontale au niveau des statocystes et comportement des statolithes.
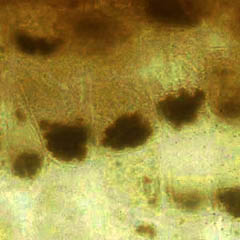
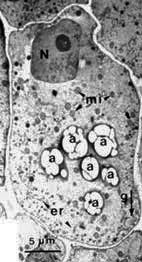
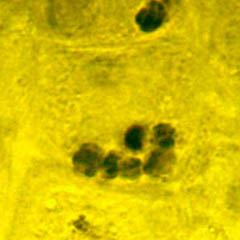
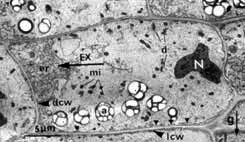
La position des statolithes peut être visialisée en microscopie photonique (coloration au lugol de l'amidon par exemple) et vérifiée en microscopie électronique (clichés G. Perbal)
 |
 |
 |
 |
|---|---|---|---|
| Amyloplastes en MO (à gauche) et MET (à droite) dans les statocystes d'une racine placée verticalement. | Amyloplastes en MO (à gauche) et MET (à droite) dans les statocystes d'une racine placée verticalement. | ||
La sédimentation des amyloplastes dans les statocytes apparait donc comme l'un des mécanismes majeurs de perception du signal gravitropique. Cependant, la réponse gravitropique, qui consiste en une courbure de la racine, suppose une modification des vitesses d'élongation de part et d'autre de l'axe de la racine. On peut dès lors s'interroger sur les mécanismes qui relient la sédimentation des amyloplastes à cette réponse physiologique. En tout premier lieu, il doit y avoir au niveau des statocytes transformation du signal physique "sédimentation des amyloplastes" en un message biochimique.
Transformation d'un signal physique en un message biochimique
Expérimentalement, l'emploi d'inhibiteurs des canaux membranaires étirement-dépendant supprime le gravitropisme. On peut dès lors supposer que la sédimentation des amyloplastes, en agissant physiquement sur ces canaux, les active et permet des modifications de concentrations de certaines substances, qui pourraient alors jouer le rôle de messagers biochimiques. On peut supposer simplement que la chute des amyloplastes sur la membrane de la face inférieure du statocyte pourrait, en étirant cette membrane, activer les canaux qu'elle contient. Une hypothèse complémentaire de la précédente suppose que les amyloplastes sont reliés au réseau de filaments d'actine (cytosquelette) existant dans le statocyte. Ces filaments d'actine seraient eux même reliés à la membrane de la cellule. Les mouvements des amyloplastes produiraient donc des tensions dans les filaments d'actine. Ces tensions se propageraient le long des filaments jusqu'à la membrane, produisant ainsi l'activation des canaux sensibles à la déformation. Quoi qu'il en soit, le passage d'un signal physique (la sédimentation des amyloplastes) à un signal biochimique semble donc se faire par le bais de l'ouverture de canaux membranaires sensibles à la déformation.
Intervention de l'auxine
 |
 |
|---|---|
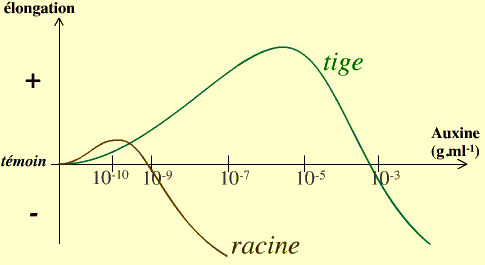
| Réponse de croissance à des concentrtions croissantes d'auxine. La sensibilité des racines et des tiges est très différentes. Les doses qui stimulent la croissance des tiges sont inhibitrices de la croissance des racines. | De la gélose contenant de l'auxine en forte concentration inhibe la croissance du côté où elle est appliquée. |
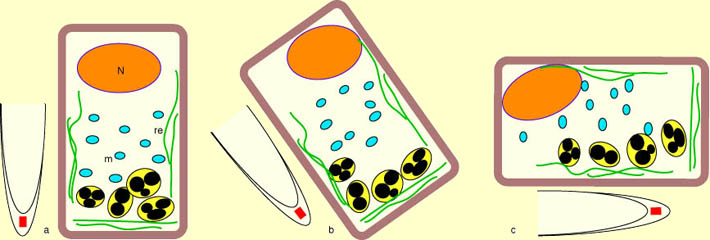
Il semble que les statotithes appuient sur la membrane plasmique (ou sur des membranes du réticulum) et provoquent une stimulation du transport de l'auxine. Le schéma suivant montre comment cette stimulation par les statolithes peut provoquer une distribution asymétrique de l'auxine.
 |
 |
L'auxine, en provenance
des jeunes tiges, circule de haut en bas dans le cylindre central
des racines puis remonte dans la région corticale de manière
symétrique mais en faible dose favorable à la croissance
des racines. |
Lorsque la racine est couchée
horizontalement, le transport d'auxine est décalé
dans le sens de la pesanteur au niveau de la coiffe. L'auxine remonte
alors avec une plus forte concentration dans la région basse
vers la zone de croissance. |
Cette différence de concentration en auxine pemettrait une croissance différentielle et donc la courbure de la racine.
 |
L'auxine étant inhibitrice
de la croissance des cellules de racine à forte concentration,
la racine se courbe vers le bas. |
Et le gravitropisme négatif des tiges ?
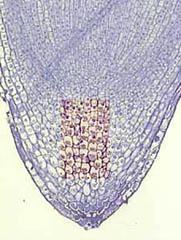
Un mécanisme de même type doit exister chez les tiges et permettre un gravitropisme négatif par suite de l'action stimulante de l'auxine aux mêmes doses sur la croissance des cellules de tige. Les lieux de réception semblent actuellement moins bien définis. Ce sont peut être les cellules de l'endoderme chez certains hypocotyles ou d'autres cellules situées à la périphérie des tissus conducteurs. Ces cellules, appelées statocytes tout comme celles contenues dans la coiffe racinaire, contiennent de volumineux grains d'amidons (statolithes) qui sédimentent sous l'effet de la pesanteur.
 |
|---|
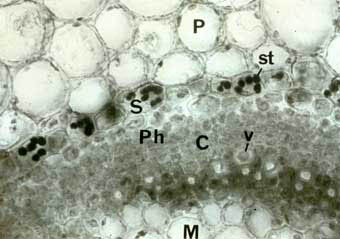
Coupe transversale dans une tige de véronique (Veronica arvensis), coloration au lugol. |
Cette sédimentation déclencherait l'activation de transporteurs membranaires actifs de l'auxine. L'action de ces transporteurs créerai un gradient d'auxine entre les face inférieures et supérieures de la tige. Les concentrations d'auxine présentes dans la tige correspondent, par opposition à ce qui existe dans la racine, à la zone où une augmentation de concentration d'auxine augmente l'élongation cellulaire. La courbure se fait de la même façon que dans la racine, par une différence d'élongation des cellules entre les faces inférieure et supérieure de la tige. Le signal responsable de cette modulation de l'élongation cellulaire serait donc ici aussi l'apparition de gradients latéraux de la concentration d'auxine consécutive à la stimulation gravitropique.
Références: les photographies en MET des statocystes sont titées de G.Perbal et D.Driss-Ecole, Microgravité et gravitropisme racinaire; Acta Botanica Gallica, 140-6, p. 615-632, 1993.