| Accueil | B.Mol | Gen | Web |


Morphologie externe- Au sortir de la période de clivage, des mouvements cellulaires de grande ampleur apparaissent et remanient les trois feuillets germinatifs : ectoderme, mésoderme et endoderme. Le blastopore apparaît sur la face dorsale dans l'hémisphère végétatif sous forme d'un sillon incurvé, appelé encoche blastoporale. En morphologie externe, la jeune gastrula de xénope ressemble à celle des autres amphibiens et permet de définir les orientations ainsi que les termes généraux suivants (fig.17).
-

Figure 17. Schéma d'une jeune gastrula en vue dorsale (A) et en vue de profil gauche (B). L'encoche blastoporale représentée en brun foncé apparaît dans l'hémisphère végetatif (HV) du côté dorsal de l'embryon. La zone marginale correspond aux territoires situés dans la région équatoriale. Dorsalement, la zone marginale comprend un territoire situé au dessus de l'encoche blastoporale, la lèvre dorsale du blastopore. HA: Hémisphère animal, HV: Hémisphère végétatif, PA: Pôle animal, PV: Pôle végétatif.
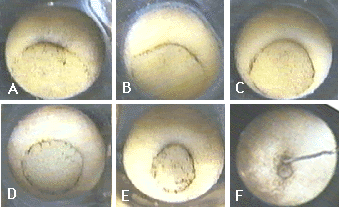
Le sillon blastoporal (fig.18 A) s'allonge en décrivant une large courbe autour du pôle végétatif (fig.18 B et C), jusqu'à former un cercle qui délimite le bouchon vitellin (fig.18 D). Ce dernier est internalisé dans l'embryon. Le diamètre du cercle blastoporal ainsi formé diminue donc progressivement (fig.18 E et F). Sur la figure 18F, le fil métallique montre le bouchon vitellin résiduel vers la fin de la gastrulation. Par commodité mnémotechnique, il est d'usage de donner des noms imagés aux différents stades de la gastrulation (fig.18 et 20). Chez le xénope, les mouvements de la gastrulation durent environ 7 à 8 heures à température ambiante.
|
|
| Figure 18. Quelques stades de la gastrulation vus par l'hémisphère végétatif. A, stade « encoche blastoporale ». B, stade « anse de panier ». C, Stade « fer à cheval ». D, « bouchon vitellin » stade jeune. E, stade « bouchon vitellin » âgé. F, stade « bouchon vitellin » final. |
Dès que l'embryon atteint le stade du bouchon vitellin, on distingue, en plus de la lèvre dorsale du blastopore, les lèvres latérales et la lèvre ventrale du blastopore devenue circulaire (fig.19).
|

|
|
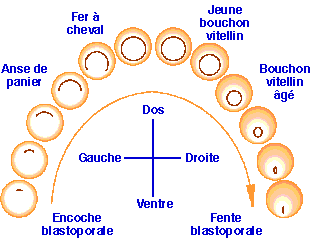
On peut résumer les différentes étapes de la gastrulation par l'évolution du blastopore depuis le stade de l'encoche blastoporale jusqu'au stade de la fente blastoporale (fig.20). Cette séquence de schémas permet de se rendre compte que les mouvements de la gastrulation sont initiés dans la région dorsale, progressent dorsolatéralement, puis ventrolatéralement pour enfin se terminer dans la région ventrale. Les mouvements gastruléens se propagent donc dans le sens dorsoventral.
|
|
| Figure 20. Représentation de la gastrulation vue par l'hémisphère végétatif montrant l'évolution du blastopore depuis sa formation (stade encoche blastoporale) jusqu'à l'achèvement de la gastrulation (stade fente blastoporale). (Voir l'animation) |
Cette transformation morphologique externe s'accompagne de mouvements cellulaires internes de très grande ampleur qui concernent tous les territoires de l'embryon.
Morphologie interne
Compte tenu de la complexité des mouvements gastruléens
dans les trois dimensions de l'espace, nous n'analyserons que les mouvements
cellulaires dans le plan médian, représentatifs de l'ensemble.
Dans la région dorsale, alors que l'encoche blastoporale apparaît
à la surface de l'embryon, les cellules internes situées
à l'angle du plancher et du toit du blastocoele adhèrent
à une structure nouvellement formée à la surface
des cellules du toit : la matrice extracellulaire (MEC). La MEC sert de
support de migration à ces cellules dites pionnières qui
se déplacent en direction du pôle animal. Elles forment un
front de migration (fig.21).
|

|
| Figure 21. Représentation schématique de la région dorsale d'une jeune gastrula à partir de quelques coupes histologiques au niveau du blastopore, du front de migration et du toit du blastocoele. |
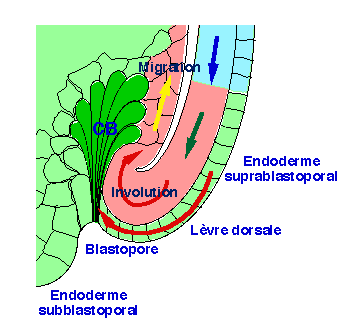
Le fond de la dépression blastoporale est formé de cellules endodermiques fortement allongées vers l'intérieur de la masse endodermique. Elles sont appelées cellules en bouteille. A la surface de l'embryon, les territoires dorsaux se dirigent vers le blastopore et entrent en contact avec les cellules en bouteille qui forment un rempart compact. Leur rôle est, semble - t'il, de forcer les territoires mésodermiques à s'enrouler sur eux mêmes (mouvement d'involution), de façon à orienter leur déplacement à la suite des premières cellules migrantes (fig.22).
|
|
| Figure 22. Un schéma d'interprétation d'une coupe sagittale dans la région dorsale rend compte des mouvements d'involution. Les territoires mésodermiques externes (rose) se dirigent vers le blastopore (flèche verte). Au contact des cellules en bouteille (vert foncé), ils s'enroulent en un mouvement d'involution (flèche rouge), s'engagent dans l'embryon à la suite des cellules mésodermiques en migration (flèche jaune). L'endoderme suprablastoporal qui constitue la couche superficielle de la lèvre dorsale du blastopore suit parallèlement l'involution du mésoderme (flèche rouge). (d'après Keller et Jansa, 1992). |
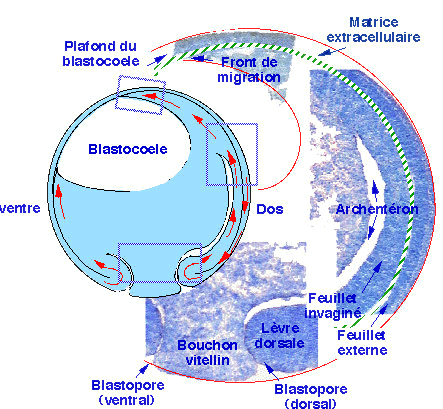
Au cours de la gastrulation, les tissus qui entrent dans le blastocoele suivent le front de migration. Simultanément, le blastopore se creuse d'une cavité nouvelle appelée cavité digestive primitive ou archentéron (fig. 23). L'archentéron se forme dans la région dorsale de l'embryon. L'ampleur des mouvements de la gastrulation est maximum dans la région dorsale et minimum dans la région ventrale. Cette observation indique que les mouvements gastruléens se déroulent selon un gradient dorsoventral d'ampleur décroissante.
|
|
| Figure 23. Représentation schématique de la région dorsale d'une gastrula âgée à partir de quelques coupes histologiques au niveau du bouchon vitellin, de la lèvre dorsale du blastopore, de l'archentéron et du front de migration. |
Mise en évidence des mouvements cellulaires
-
- Les mouvements cellulaires de la gastrulation ont été mis en évidence pour la première fois par l'embryologiste allemand W. Vogt en 1929 chez deux espèces d'amphibien, un triton et un crapaud. Il eut l'idée de poser sur la surface de l'embryon, des fragments d'agar solidifiés et imprégnés de colorants vitaux tels que le rouge neutre ou le bleu de Nil. En diffusant dans l'eau, le colorant marque les cellules au contact de l'agar en quelques minutes. Il suffit alors d' ôter le fragment d'agar et d'observer l'évolution des cellules marquées parmi toutes les autres cellules non colorées (fig.24). Plus tard, l'opération a été effectuée chez le xénope par l'embryologiste américain R. Keller en 1975 et 1976.
-
-
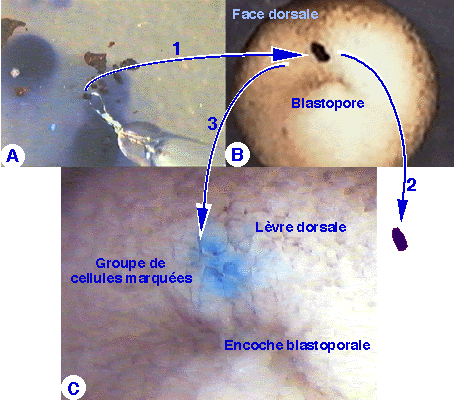
Figure 24. Marquage coloré sur un embryon de xénope au stade jeune gastrula. En A, des fragments d'agar fortement imprégnés de bleu de Nil ont été éparpillés sur le fond d'une coupelle remplie d'eau. A l'aide d'une baguette de verre terminée par une anse métallique, un fragment de quelques dizaines de microns est saisi, transporté (1) puis appliqué à la surface de l'embryon (B). Environ 5 minutes après cette opération, le fragment d'agar est retiré (2). Le résultat est la coloration d'un groupe de cellules (3) à l'emplacement du fragment d'agar. Dans cet exemple, les cellules marquées sont situées dans le territoire de la lèvre dorsale du blastopore (C).
De cette façon on conçoit que, de proche en proche, on
puisse marquer toute la surface de l'embryon par groupes de quelques cellules.
Les résultats sont doubles.
Premièrement, les marques colorées peuvent être retrouvées
plusieurs jours plus tard dans les ébauches d'organes de l'embryon.
On peut donc connaître, avec une certaine précision, la destinée
des cellules marquées dès le début de la gastrulation
et dresser ainsi une carte des destins cellulaires (carte des territoires
présomptifs) (fig.25).
|
|
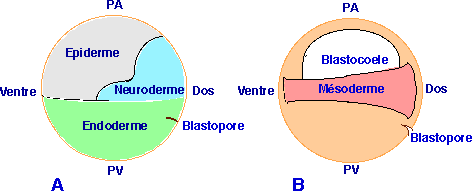
| Figure 25. Carte des territoires présomptifs de xénope au tout début de la gastrulation (d'après Keller, 1976). En A et B, sont représentés respectivement les tissus superficiels et profonds. On remarque que les tissus mésodermiques situés dans la zone marginale sont exclusivement constitués de cellules profondes. |
Deuxièmement, le déplacement des marques colorées
nous renseigne sur les mouvements d'ensemble qu'effectuent ces territoires
embryonnaires pendant la gastrulation. Des territoires se redéploient
à l'intérieur de l'embryon en comblant l'espace interne
formé par le blastocoele. C'est l'invagination. C'est le
cas des territoires mésodermiques et endodermiques. Simultanément,
les autres territoires s'étalent à la surface de l'embryon
et le recouvrent entièrement. On parle de mouvements d'épibolie.
C'est le cas des territoires ectodermiques (épiderme et neuroderme).
Du point de vue de la mécanistique cellulaire, on distingue les
mouvements d'intercalation radiale qui contribuent à la
progression des mouvements d'épibolie et concernent par conséquent
les territoires ectodermiques ; les mouvements de migration cellulaire
sur le toit du blastocoele, moteur de l'invagination du mésoderme
antérieur et ventral ; les mouvements de convergence- extension,
moteur de l'invagination du mésoderme dorsal et de l'allongement
antéropostérieur de l'embryon.
L'ensemble de ces mouvements est commun au développement embryonnaire
des amphibiens ayant fait l'objet d'études embryologiques. Cependant,
le xénope présente deux particularités au niveau
des mouvements d'épibolie et d'invagination.
Chez le xenope, à la fin de la segmentation, la région
de l'hémisphère animal à l'origine de l'ectoderme
est composée d'un épithélium
tristratifié formé d'une assise cellulaire superficielle
et de deux assises cellulaires profondes. Dès le début de
la gastrulation, les deux assises de cellules profondes s'interpénètrent
à la manière d'une fermeture Eclair (fig.26).
Ce mécanisme progresse du pôle animal vers la zone marginale.
En fin de gastrulation, la couche profonde n'est donc plus formée
que d'une seule assise de cellules. En conséquence, alors qu'au
stade blastula, l'ectoderme comprend trois couches de cellules, au stade
gastrula âgée, le même feuillet n'est plus formé
que par deux couches de cellules, une assise superficielle et une profonde.
La résolution en une seule couche cellulaire d'une assise originelle
bistratifiée entraîne nécessairement une augmentation
de surface. Parallèlement, la couche de cellules superficielles,
bien que ne participant pas aux mouvements d'intercalation radiale, s'aplatit
et s'étale en suivant cette augmentation de surface. Au total,
les mouvements d'intercalation radiale des cellules profondes ainsi que
l'étalement des cellules superficielles rendent compte du mouvement
d'épibolie qui oeuvre au recouvrement de l'ensemble de l'embryon
par le feuillet ectodermique.
|
|
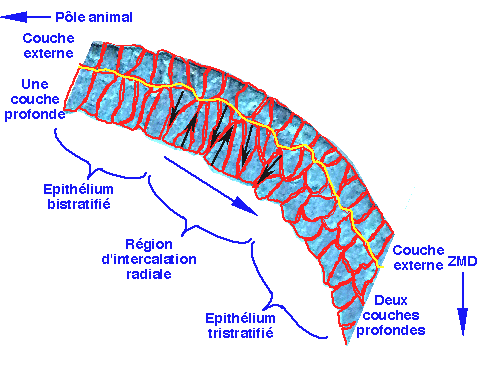
| Figure 26. Fragment dorsal d'une coupe méridienne de l'hémisphère animal. Les cellules ont été détourées en rouge pour mettre en évidence l'organisation cellulaire. La ligne jaune représente la limite entre les cellules superficielles et les cellules profondes. Les cellules profondes s'intercalent entre elles (flèches noires). Le mouvement d'intercalation radiale progresse depuis le pôle animal vers la zone marginale dorsale (ZMD). |
Migration
Dans le cas de la gastrulation des amphibiens, on entend par migration
cellulaire, le déplacement d'une cellule d'un point à un
autre à l'aide d'un support de migration extracellulaire appelé
matrice extracellulaire (MEC). Chez le xenope, les mouvements de migration
cellulaire concernent les premiers territoires mésodermiques qui
entrent dans le blastocoele et qui sont à l'origine du mésenchyme
céphalique, du coeur, des îlots sanguins, et des lames latérales.
Le reste du mésoderme à l'origine de la chorde et des somites
involue plus tardivement selon un mécanisme différent appelé
convergence- extension.
Il est admis que, si les mouvements de migration cellulaire existent lors
de la gastrulation du xenope, ceux-ci ne constituent pas le moteur prépondérant
de l'invagination. Des expériences ont même montré
que la délétion du toit du blastocoèle ou l'inhibition
de l'adhérence cellulaire à la MEC n'empêchent pas
les mouvements d'invagination et la fermeture du bouchon vitellin. Par
contre, il en va tout autrement de la gastrulation dans les embryons de
triton où la migration cellulaire est considérée
comme essentielle .
En conséquence, l'étude de la migration cellulaire ne sera
pas abordée ici. Elle fera l'objet d'un examen ultérieur
plus détaillé chez une autre espèce d'amphibien où
elle est indispensable.
Convergence - extension
Les mouvements de convergence- extension concernent les territoires dorsaux et latérodorsaux de la gastrula, c'est à dire le mésoderme de la chorde, des somites ainsi qu'une partie du neuroderme. Ils consistent en une intercalation cellulaire médiolatérale. Les cellules disposées latéralement par rapport au plan médian convergent vers celui-ci. Elles sont donc obligées de s'intercaler avec leurs voisines et provoquent une pression des tissus sur la ligne dorsale de l'embryon. La résultante de ces mouvements est un allongement antéropostérieur des territoires concernés. Des expériences de lignage cellulaire utilisant des marqueurs fluorescents ainsi que la vidéomicroscopie ont permis de démontrer l'existence de ces mouvements (fig. 27).
|
|
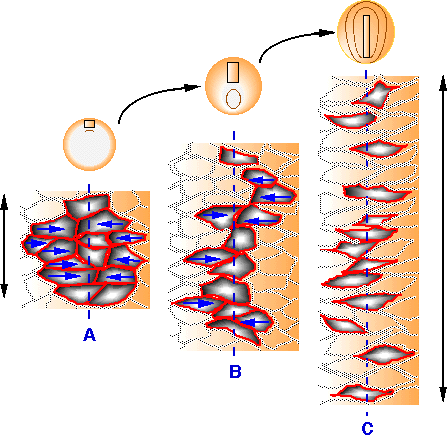
| Figure 27. Représentation schématique des mouvements de convergence-extension dans la zone marginale dorsale. Un groupe de 13 cellules parmi l'ensemble des cellules du territoire dorsal (grisé) est suivi depuis le début de la gastrulation jusqu'au début de la neurulation (A à C). Le plan médian est représenté par la ligne bleue discontinue. Au stade jeune gastrula, les cellules forment un amas cohésif (A). Les cellules les plus latérales de ce groupe deviennent bipolaires et se dirigent vers le plan médian. Elles sont convergentes (flèches bleues). Au cours de la gastrulation, ces mêmes cellules s'intercalent avec leurs voisines et au stade gastrula âgée la forme du groupe de cellules a changé. Il s'est s'allongé le long du plan médian (B). Au début du stade neurula, le phènomène s'est considérablement accentué. Les cellules se sont espacées et alignées le long du plan médian (C). Le groupe de cellules initial a donc subi une extension antéropostérieure (doubles flèches noires) (D'après Shih et Keller, 1992). |
Les mouvements d'extension ont pour effet d'étendre les tissus dorsaux et dorsolatéraux vers la région postérieure de l'embryon, c'est à dire vers le blastopore. Combinés avec les mouvements d'involution, la pression exercée par les mouvements d'extension provoque le recul du bord d'enroulement du blastopore vers le pôle végétatif. Les tissus dorsaux et latérodorsaux recouvrent donc le bouchon vitellin (fig.28 et 29).
|

|
| Figure 28. Gastrulation vue par l'hémisphère végétatif. Représentation schématique de la progression des territoires dorsaux et dorsolatéraux vers le blastopore. |
|
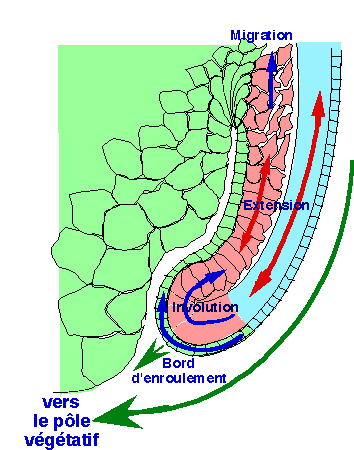
|
| Figure 29. Un schéma d'interprètation d'une coupe sagittale dans la région dorsale rend compte des mouvements d'extension. Au cours de la gastrulation, les mouvements de convergence provoquent l'extension (flèches rouges) du mésoderme invaginé (rose) ainsi que du neurectoderme externe (bleu clair). En conséquence, le bord d'enroulement progresse vers le pôle végétatif et recouvre l'endoderme subblastoporal (flèches vertes). (d'après Keller et Jansa, 1992). |
Conclusion
Au stade de la gastrulation, des mouvements cellulaires de grande ampleur affectent l'ensemble de l'embryon. Des remaniements cellulaires en découlent et redistribuent les tissus embryonnaires. Les trois feuillets originaux, ectoderme, mésoderme et endoderme sont maintenant organisés de manière concentrique : l'endoderme profond, l'ectoderme superficiel et le mésoderme en position intermédiaire (fig. 30, 31). Une nouvelle cavité est formée au détriment du blastocoele : l'archentéron.
 |
 |
| Figure 30. Schéma d'une coupe transversale au début de la gastrulation montrant l'organisation étagée de l'ectoderme, du mésoderme et de l'endoderme. | Figure 31. Schéma d'une coupe transversale à la fin de la gastrulation montrant l'organisation concentrique des feuillets embryonnaires. |
L'environnement de chaque cellule a donc changé, ce qui génère de nouveaux contacts entre les tissus. Ainsi, au cours de l'invagination, des contacts qui s'établissent entre le mésoderme dorsal et l'ectoderme sont une des composantes de l'induction du neurectoderme. Le modelage de l'embryon peut alors commencer avec l'apparition des ébauches d'organes. Le premier étant le système nerveux, l'embryon accède au stade neurula.
Références citées :
- Keller, R. E. (1975). Vital dye mapping of the gastrula and neurula of Xenopus laevis. I. Prospective areas and morphogenetic movements in the superficial layer. Dev. Biol. 42, 222-241.
- Keller, R. E. (1976). Vital dye mapping of the gastrula and neurula of Xenopus laevis. II. Prospective areas and morphogenetic movements in the deep layer. Dev. Biol. 51, 118-137.
- Keller, R.E. and Jansa, S. (1992). Xenopus gastrulation without a blastocoel roof. Dev.Dyn. 195, 162-176.
- Shih, J. and Keller R.E. (1992) Cell motility driving mediolateral intercalation in explants of Xenopus laevis. Development 116, 901-914.
- Vogt,W. (1929). Gestalunganalyse am Amphibienkeim mit örtlicher vitalfärbung. II. Teil. Gastrulation und Mesodermbildung bei Urodelen und Anuren. Whilhem Roux Arch. Entwick. Mech. Org. 120, 384-706.
