| Accueil | B.Mol | Gen | Web |


-
1) Contrôle par le système nerveux autonome
-
L’état plus ou moins contracté
des différentes CML de l’organisme peut être contrôlé
à la fois par le système nerveux et par le système
hormonal. Selon l’organe, le tissu, le type cellulaire ou le
territoire considéré, le contrôle est différent
même si certaines hormones exercent le même effet sur
les CML quelles que soient leurs fonctions. Le CGRP par exemple
exerce une action de relaxation aussi bien sur les CML vasculaires
que sur les CML utérines ou entériques.
Les modes d’actions du système nerveux et du système
hormonal sont différents, de part la structure du vaisseau,
et de part la localisation des CML dans ces vaisseaux au niveau
de la média. Effectivement, si les afférences nerveuses
peuvent être en contact avec les CMLV, les hormones sont transportées
par le sang et ne peuvent donc pas être en contact direct
avec les CMLV. Même dans les gros vaisseaux, au niveau du
vaso vasorum, il persiste une couche de cellules endothéliales
qui limite l’accès des hormones aux CML. Les hormones
agissent donc soit par diffusion, soit par effet indirect via les
cellules endothéliales. Cependant, les substances libérées
par les terminaisons nerveuses peuvent également diffuser
vers les cellules endothéliales si elles ne sont pas dégradées
ou recaptées immédiatement. (Fig. 8) Dans ce cas,
ces neurotransmetteurs vont agir comme des hormones paracrines.
Ainsi, la limite entre hormones et neurotransmetteurs devient plus
floue.
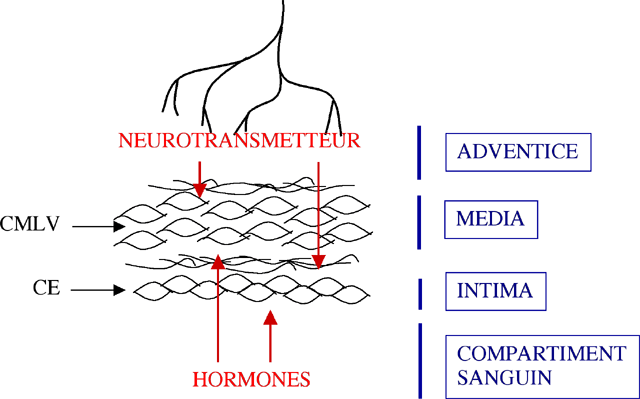 |
| Figure 8 : Mode d’action des hormones et des neurotransmetteurs sur les CMLV et sur les cellules endothéliales (CE). La biodisponibilité et l’abondance des neurotransmetteurs et des hormones déterminent leur cible cellulaire, soit les CMLV, soit les cellules endothéliales (CE). |
Le système nerveux autonome contrôle l’activité
des différents organes. Il n’est pas séparé
du système nerveux central, mais agit de façon autonome
sur des organes tels que le cœur, les vaisseaux, le système
digestif et bien d’autres. Le système nerveux autonome
est divisé en deux parties : le système nerveux sympathique
et le système nerveux parasympathique. Ces deux systèmes
sont différenciés par le neurotransmetteur post- ganglionnaire
libéré lors d’une stimulation : noradrénaline
dans le cas du système sympathique et acétylcholine
pour le système parasympathique.
Beaucoup d’organes sont innervés à la fois par
le système sympathique et par le système parasympathique,
et c’est le cas des vaisseaux et donc des CMLV. Il faut préciser
que la séparation anatomique des systèmes nerveux
sympathique et parasympathique ne cadre pas toujours avec la réalité.
La présence de fibres cholinergiques dans les troncs sympathiques
a souvent été démontrée (Dale et Feldberg,
1934), de même que la présence de fibres adrénergiques
dans les troncs nerveux généralement considérés
comme parasympathiques.
Les fibres sympathiques participent au tonus vasculaire et ont
une influence vasoconstrictrice. L’existence de fibres vasoconstrictrices
fut mise en évidence dès 1851 par Claude Bernard.
La noradrénaline libérée au niveau des fibres
post-ganglionnaires sympathiques peut agir sur des récepteurs
α-adrénergiques des CMLV et provoquer une vasoconstriction.
La section de l’innervation sympathique ou l’administration
d'α-bloquant provoque une augmentation du flux sanguin significative
: à débit constant, c.a.d activité cardiaque
constante en terme de force et de fréquence, la vitesse augmente
si le calibre diminue. La vasodilatation induite par une baisse
de l’activité des fibres sympathiques est importante
et participe à la vasodilatation autant que l’activation
du système antagoniste : le système parasympathique.
A pression artérielle normale (125 mmHg/75 mmHg) les vaisseaux
sont sous " tonus sympathique " : c’est le système
sympathique qui est en activité.
L’action de la noradrénaline sur des récepteurs
de type β-adrénergique provoquerait au contraire une
vasodilatation. C’est donc l’expression de l’un ou
de l’autre type de récepteurs qui détermine la
réponse de la CML au système sympathique. Cette distinction
pharmacologique des récepteurs adrénergiques a été
mise en évidence par Ahlquist dès 1948 (Ahlquist,
1948).
1.1.1 Récepteurs alpha adrénergiques
Il existe deux types de récepteurs α-adrénergiques,
les récepteurs α1 et les récepteurs α2,
distingués pharmacologiquement selon leurs affinités
pour divers agonistes et antagonistes. Les travaux de biologie moléculaire
ont démontré que les récepteurs α1 étaient
couplés à une production d’IP3 via l’activation
d’une PLC par une protéine Gq/11.(Guimaraes et Moura,
2001). On retrouve les récepteurs α1 notamment sur
les artères irriguant les reins, les territoires splanchniques
et la peau. Dans ces organes, la fixation des catécholamines
provoque une vasoconstriction et donc une diminution du débit
sanguin local.
Les récepteurs α2 se retrouvent quant à eux
essentiellement au niveau des veines où leur activation provoque
également une faible vasoconstriction qui favorise le retour
veineux. Ils agiraient en activant une protéine Gi, qui inhiberait
l’activité des adenylyl cyclase (AC), mais aussi par
inhibition de l’ouverture des CCVDs, et activation des canaux
potassiques (Guimaraes et Moura, 2001).
1.1.2 Récepteurs béta adrénergiques
Comme pour les récepteurs alpha, il existe plusieurs isoformes de récepteurs β-adrénergiques : les récepteurs β1, β2 et β3 (Lands 1967, Bylund et al., 1994). On retrouve les récepteurs ß1 au niveau du cœur et des artères coronaires où la fixation des catécholamines provoque une dilatation des artères coronaires et au niveau du cœur un effet ionotrope. On retrouve les récepteurs β2 au niveau des artères des muscles squelettiques. Tous les récepteurs β-adrénergiques sont couplés à une protéine Gs qui va activer la production d’AMPc par une AC (Guimaraes et Moura, 2001). Dans certains cas cependant, les récepteurs β-adrénergiques sont couplés à une protéine Gi qui va inhiber la production d’AMPc dans les cellules.
Les fibres parasympathiques ont une influence vasodilatatrice.
Leur rôle est moins important que celui des fibres sympathiques
puisque à l’état basal, l’organisme est
sous tonus sympathique. C’est encore Claude Bernard qui démontra
dès 1858 l’existence d’une vasodilatation induite
par la stimulation d’un nerf.
1.2.1 Fibres cholinergiques
Le système nerveux parasympathique cholinergique innerve principalement au niveau du système vasculaire les artères cérébrales et les artères coronaires. Ces fibres ne sont pas spontanément actives, mais vont libérer de l’acétylcholine (Ach) en cas de stimulation. L’Ach agit ensuite au niveau des récepteurs muscariniques sur la CMLV mais aussi sur les cellules endothéliales. Sur les CMLV, l’activation de récepteurs muscariniques de type M3 (Goyal, 1989) induit une élévation du Ca2+ cytoplasmique consécutive à l’activation d’une PLC, qui va aboutir à une contraction de la cellule. Cependant, au niveau des cellules endothéliales, l’Ach provoque une libération de NO, aboutissant à la vasorelaxation. L’Ach en injection intraveineuse provoque ainsi une baisse de la pression artérielle par vasodilatation nette. L’effet des afférences parasympathiques est moins univoque : il serait vasodilatateur dans le cas où l’épaisseur de la média est faible ou lorsque la libération d’Ach est abondante, permettant la diffusion de l’Ach vers les cellules endothéliales. Cet effet vasodilatateur pourrait aussi être dû à une diminution de la libération de noradrénaline au niveau des terminaisons sympathiques (par effet sur des récepteurs muscariniques pré-sympathiques). Sur les CML non vasculaires comme, par exemple, les CML gastro-intestinales ou bronchiques, l’Ach a des effets clairement constricteurs. C’est donc la présence des cellules endothéliales qui est responsable de son effet vasodilatateur.
1.2.2 Autres neurotransmetteurs
Les autres neurotransmetteurs du système parasympathique sont le VIP, l’histamine, les purines, la sérotonine, la substance P, le CGRP et les enképhalines. Toutes ces substances peuvent être libérées par le système nerveux sympathique ou parasympathique en même temps ou non que la noradrénaline ou que l’Ach.
Le système nerveux sensoriel peut être impliqué
dans le contrôle de l’état contractile des CML
des vaisseaux cutanés par un mécanisme appelé
réflexe d’axone et décrit dès 1923 par
Bayliss. Il est facile de constater chez l’homme que des stimuli
divers portés sur la peau provoquent l’apparition d’une
vasodilatation limitée au territoire excité et à
son voisinage immédiat. Cette vasodilatation est visible
par la rougeur et la chaleur qui l’accompagnent. Les nerfs
nociceptifs de type Aδ et C vont en périphérie
se diviser en deux branches : une branche sensitive et une branche
artériolaire., L’influx nerveux, à la suite d’un
stimulus douloureux, va se propager le long de la fibre dans le
sens centripète, emprunter à partir du point de ramification
une fibre collatérale parcourue alors dans le sens antidromique
vers la périphérie jusqu'à la musculature artériolaire.
La fibre nerveuse va libérer à ce niveau la substance
P et le CGRP, responsable de la vasodilatation mais également
d’une hypersensibilité (Fig. 9). La zone lésée
va être plus sensible, afin de la préserver d’une
autre atteinte.
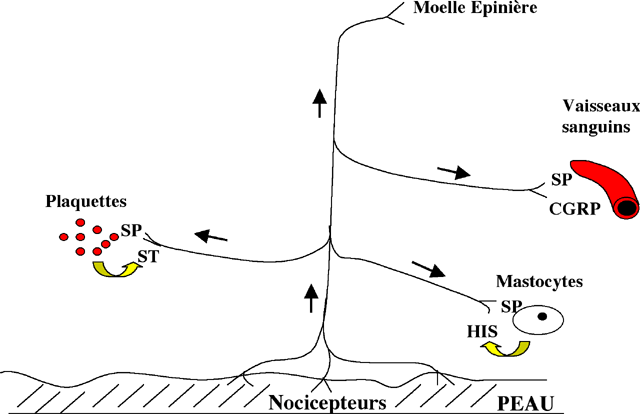 |
| Figure 9 : Réflexe d’axone dans les voies nociceptives. Un simulus douloureux va provoquer l’activation des voies nociceptives. L’influx nerveux peut se propager dans le sens antidromique et libérer localement la substance P (SP) et le CGRP. Cette libération va provoquer la libération par les plaquettes de sérotonine (ST) et d’histamine (HIS) par les mastocytes. Le CGRP et l’histamine libérés sont responsables de la vasodilatation locale |
| A- Localisation, structure
et fonction des CMLV B- Mécanisme de la relaxation/contraction de la CMLV C- Régulation de la contraction/relaxation de la CMLV Bibliographie |
1) Contrôle
par le système nerveux autonome 2 ) Contrôle hormonal 3) Autres contrôles |
