| Accueil | B.Mol | Gen | Web | sommaire |

Modalités de la digestion : les glandes sécrétrices d'enzymes digestives
Abréviations :
sp. : espèce (du latin species)
sp. pl. : plusieurs espèces
Ce chapitre sur la digestion ne vise pas à l'exhaustivité. Il reprend beaucoup des données citées par Juniper et collègues (voir bibliographie).
Certaines espèces ne figurent pas dans ce chapitre sur la digestion, comme les Utricularia.
Rappel d’éléments du métabolisme de l’azote chez les végétaux
Ce qu'il faut retenir c'est que d'une manière générale la source d'azote des plantes se présente sous forme d'ion nitrique, mais elles peuvent assimiler l'azote sous forme d'ion nitreux ou ammoniacal. Remarquez aussi que si la voie d'absorption classique passe par les racines, on utilise l'aspersion du feuillage, par exemple des fruitiers, avec des solutions nutritives ce qui montre que de nombreux végétaux peuvent absorber des ions nitriques ou ammoniacaux par les feuilles.
De plus les végétaux sont capables d'assimiler l'azote sous forme organique de faible poids moléculaire: acides aminés surtout acides aspartiques et glutamique, asparagine, glutamine, urée, acide urique…
Dans la plante, l'azote nitrique est réduit en azote ammoniacal, (NH4+). Cet azote ammoniacal est immédiatement incorporé sous forme d'acides aminés ou bien incorporés sous forme d'amide comme les acides aminés asparagine et glutamine.
Les plantes carnivores vivent dans des stations naturellement dépourvues en source d'azote assimilable. Elles sont dépourvues d'organisme symbiotique fixateur d'azote contrairement aux Légumineuses, aux Aulnes ou aux Casuarina.
Comment l'azote des proies est-elle transférée dans la plante carnivore?
Très précocement, les scientifiques d'origine européenne ont observé la transformation des proies, comme celle de petits morceaux de viande par les plantes carnivores, suggérant l'existence d'une digestion comparable à celle des animaux. (Arthur Dobbs découvre la Dionée en 1759 et l'appelle "Sensitive piège à mouche". Ellis la décrit aux alentours de 1770, il la considère comme une plante carnivore. Darwin, cite des observations de digestion antérieurement à 1875, chez d'autres plantes carnivores). Il n'y a pas lieu de croire que les amérindiens et les populations d'Asie aient pu ignorer de tels phénomènes, mais je n'ai pas de bibliographie à ce sujet.
Toutes les modalités de la digestion ne sont pas connus ou bien sont imparfaitement connu, la digestion chez les Utriculaires en est un exemple. La présentation qui suit en présentent quelques aspects mais ne vise pas à l'exhaustivité.
Pour qu'il y ait digestion, il faut un contact étroit entre les enzymes responsables de celle-ci et leur substrat. Dans ces conditions, le meilleur contact a lieu dans une phase liquide. Il est donc nécessaire que la proie puisse être immergée au moins partiellement dans une soupe enzymatique.
Première étape : la présence de liquide.
La proie doit être dans un liquide. Parmi les cas qui peuvent se présenter :
il est produit une fois le piège de la Dionée refermé, et après formation d'une poche étanche par le rapprochement des bords du piège.
Chez Pinguicula, les glandes pédicellées piégent la proie, mais ce sont les glandes sessiles qui contribuent à fournir le liquide et les enzymes.
Chez la Dionée la seule stimulation des poils sensibles peut amener à un efflux d'une solution ne contenant pas d'enzyme. Chez Darlingtonia, qui n'a pas de glande digestive, l'ajout d'un bouillon de boeuf au contenu de l'urne déclenche une augmentation du volume de liquide de l'urne.
Ceci ne doit pas faire oublier que la sécrétion d'enzymes et l'efflux de liquide sont souvent liés.
Le pH des gouttelettes de mucilage avant stimulation est souvent acide. Le pH du mucilage des Drosera varie de 3 à 5 suivant les espèces. De même certaines espèces de Drosera voient un abaissement du pH de 2 unités après stimulation des tentacules. Chez des Pinguicula le pH est de l'ordre de 5 et s'abaisse à pH 3 après stimulation des feuilles par une proie.
Deuxième étape : la présence d'enzymes, les conditions de leur éventuelle sécrétion par la plante.
Pour transformer, à la température ambiante, la matière organique de la proie en molécules suffisamment petites pour être réabsorbées il faut l'action d'enzymes. D'une manière générale les enzymes à l'oeuvre ont pour caractéristique une certaine stabilité ce qui leur permet d'avoir une activité étalée dans le temps. Elles ont aussi une faible spécificité vis à vis de leurs substrats. Ceci est un avantage car un organisme est un mélange de substrats fort divers et la synthèse d'un grand nombre d'enzymes spécifiques aurait un coût énergétique très élevé pour ces espèces.
L'apport d'enzymes par l'extérieur.
Il existe dans l'air et sur les proies, dans le système digestif de celles-ci des bactéries capables d'hydrolyser la matière organique. Elles sont présentes dans l'air et sur les proies. Ceci complique l'étude de l'origine des enzymes digestives. Pour être sûr que les enzymes sont bien secrétées par les plantes carnivores il faut avoir recours, en général, à des cultures axéniques (sans germe). Dans certain cas particuliers, on peut recueillir aseptiquement le contenu liquide des espèces à urnes avant que l'opercule qui clôt ces dernières ne s’ouvre.
L'existence de bactéries dans le milieu extérieur, qui peuvent hydrolyser la matière organique, est un avantage certain pour les espèces qui ne secrètent pas d'enzyme.
Parmi ces espèces on peut citer :
Certaines espèces sont dépourvues d'enzymes habituellement présentes : par exemple les Sarracenia sp. pl. sont dépourvues d'estérase et d'uréase.
Suivant les genres et les espèces, et souvent le type de glande de l’espèce considérée, la nature de ces enzymes varie. La chitinase permettrait de digérer l’exosquelette des insectes. Elle est très souvent absente. Lorsqu'elle est présente, il est difficile de dire si elle est secrétée par la plante ou bien si c'est un apport microbien. Curieusement, même quand elle est présente, dans les pièges, on constate, à l'issue de la digestion la présence des restes chitineux des insectes.
Espèces |
Type de glande |
Enzymes |
||||
estérase |
phosphatase acide |
amylase |
protéase |
ribonucléase |
||
Cephalotus folicularis |
grande |
- |
+ |
? |
- |
? |
Cephalotus folicularis |
petite |
+ |
- |
? |
+ |
+ |
Dionaea muscipula |
+ |
+ |
? |
++ |
++ |
|
Drosera capensis |
pédicellée |
++ |
+ |
? |
? |
? |
Drosera rotundifolia |
pédicellée |
+ |
++ |
? |
? |
? |
Drosophyllum |
pédicellée |
+ |
+ |
? |
++ |
+ |
Drosophyllum |
sessile |
+ |
+ |
? |
+ |
+ |
Pinguicula grandiflora |
pédicellée |
+ |
+ |
+ |
+ |
+ |
| Pinguicula grandiflora | sessile | + + |
+ + |
- |
+ + |
+ + |
Pinguicula moranensis |
sessile |
+ |
+ |
? |
? |
? |
Pinguicula vulgaris |
pédicellée |
+ |
+ |
+ |
+ |
+ |
Pinguicula vulgaris |
Sessile |
+ |
+ |
+ |
+ |
+ |
Utricularia neglecta |
quadrifide |
? |
+ |
? |
+ |
? |
Nepenthes maxima |
+ |
+ |
? |
+ |
+ |
|
Sarracenia purpurea |
+ |
+ |
? |
+ |
? |
|
Légende |
||||||
Types d’enzymes
localisés cytologiquement dans les glandes digestives.
Tableau simplifié d’après Juniper et al. (1989). |
||||||
Les proies sont, en général, de petite taille : des insectes en majorité. Les plantes absorbent les produits de digestion des protéines mais ne dégradent pratiquement pas la chitine, élément essentiel du tégument et des ailes d'insectes. On trouvera ainsi, après la digestion, une carcasse vidée de son contenu. La plante conserve les débris jusqu'à l'assèchement de la feuille ou bien, le vent et la pluie lavent les déchets.
L'apport d'enzymes issu du métabolisme de la plante
La sécrétion d'enzyme en continu
Des espèces comme les Nepenthes secrètent en continu des enzymes.
La sécrétion d'enzyme à la demande.
Exemple des Pinguicula et Dionaea.
Une manière d'économiser de l'énergie pour la synthèse d'enzymes est de n'en délivrer qu'en présence de la proie (glandes sessiles des Pinguicula et Dionaea). Ceci est d'autant plus important pour les espèces qui ne disposent pas de poche digestive permanente. Chez Pinguicula les glandes pédicellées secrètent de l'amylase.
Comment les glandes sessiles de ces espèces perçoivent la présence de la proie à digérer?
La sécrétion d'enzymes est déclenchée par le contact notamment de solutés de chlorure de sodium, d'ammoniaque d'acide urique, d'acides aminés ou de phosphates avec les glandes du piège. On ne sait pas si la stimulation d'émission de liquide par le chlorure de sodium (présent dans l'haemolymphe des insectes) déclenche aussi la sécrétion d'enzymes.
L'acide urique est un des composants principaux de l'urine des insectes. La détection de sa présence au contact des glandes des plantes carnivores augmente la probabilité que le coût de la sécrétion d'enzymes sera compensé par l'apport de nutriments provenant de la proie.
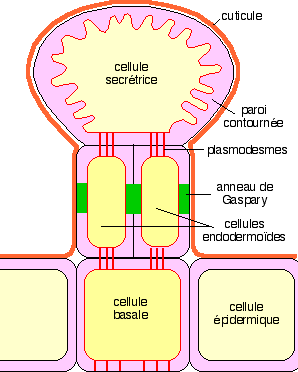 |
Schéma d'une glande sécrétrice
Variations : elles portent sur la position de la glande par rapport aux tissus du piége. Il peut exister un pédicelle vascularisé au sommet duquel s'insère la glande (cf. Drosera) ; elles portent sur le nombre de cellules sécrétrices, sur la structure de la cuticule et aussi sur la ou les cellules basales qui peuvent aussi ne pas exister, la glande se rattachant alors au système vasculaire de la plante.
Commentaires : à maturité, les enzymes s'accumulent dans des vacuoles et dans les parois de la cellule sécrétrice. Lors de la stimulation, les enzymes peuvent traverser le plasmalemme ou bien le plasmalemme se détruit et le contenu cellulaire est déversé à l'extérieur à travers les discontinuités de la cuticule.
Les plantes terrestres sont recouvertes d'une cuticule imperméable qui est un obstacle à la sortie de l'eau mais aussi des enzymes lorsqu'elles atteignent la paroi des cellules.
Chez Drosophyllum, des Drosera, des Pinguicula et des Utricularia, mais aussi chez la Dionée où ce phénomène n'apparaît qu'après la perception des stimulus chimiques, on observe au niveau des glandes digestives l'interruption de la couche cuticulaire (observations de Darwin chez la Dionée, 1875).
Chez les Utricularia chaque piége peut fonctionner plusieurs fois et réaliser des cycles de captures et digestions. Il en est de même chez la Dionaea.
Chez les Pinguicula, seule la partie de la feuille en contact avec la proie subit des transformations. Les glandes digestives, une fois leur contenu enzymatique vidé, ne recommencent pas un cycle de synthèse contrairement à Dionaea.
Une fois la digestion commencée comment les produits de celle ci sont ils réabsorbés?
Chez les Nepenthes la réabsorption des produits de la digestion se fait dans la phase de sénescence du piège.
Chez les espèces à piège à glue, ou chez la Dionaea, les produits de la digestion empruntent précocement le trajet inverse à celui de la sécrétion d'enzymes. Le marquage de composés protéiques a permis de suivre le trajet des produits de la digestion. Il a pu être constaté qu'ils empruntent les vaisseaux du xylème.
Où vont les produits de la digestion? Part du régime carnivore dans la nutrition des plantes carnivores.
La pauvreté en azote des milieux où croissent les plantes carnivores a suggéré depuis longtemps le rôle des proies dans l'apport en azote assimilable et aussi en phosphate.
Chez Dionaea : des études utilisant des acides aminés marqués au 14C ont montré que ceux ci étaient métabolisés par la plante [Rea, 1982]. Des études fondées sur l'inégale répartition de 15N dans le sol et dans les proies capturées a permis d'évaluer que celles ci fournissent 75% de l'azote de la plante. [(Schultze 2001)]
Chez Drosera erythrorhiza, des acides aminés marqués ont permis de détecter les lieux de stockage de ces produits. Lors de la pleine activité on retrouve 27 % du marquage dans les feuilles et 46% dans les tubercules destinés à passer la mauvaise saison. [Dixon 1980, cité par Juniper]
Les proies sont aussi d'une manière moindre une source de carbone.
L'analyse de la composition isotopique de l'azote du sol, d'espèces de plantes carnivores et de leurs proies a permis de déterminer la contribution de l'apport d'azote par les insectes par rapport à l'azote total de la plante.
Types de pièges |
Espèces |
Part de l'azote provenant d'insectes en % |
Piège à succion |
Utricularia vulgaris |
51,8 |
"Papier tue-mouche" |
Drosera rotundifolia |
26,5 |
Urnes |
Cephalotus follicularis |
26,1 |
Urnes |
Darlingtonia californica |
76,4 |
Urnes |
Heliamphora sp. |
79,3 |
Urnes |
Brocchinia reducta |
59,8 |
Urnes |
Sarracenia purpurea |
10 |
D'après A. M. Ellison et N. J. Gotelli, 2001, simplifié. |
||
Les expériences sur certaines espèces de plantes carnivores suggèrent que ce régime carnivore n'a pas d'effet sur le contenu de celles-ci en calcium, magnésium et potassium (Sarracenia flava). Par contre, les individus appartenant à l'espèce Triphyophyllum sp. (famille des Dioncophyllaceae) peuvent accumuler du potassium sur des sols déficients en cet élément.
En conditions limitantes en azote assimilable et en phosphate, la capture des proies permet une meilleure croissance et une meilleure production de graines.
Chez Pinguicula vulgaris, même sur un milieu normalement pourvu en éléments minéraux, la capture de proies améliore l'incorporation d'azote et de phosphate. C'est de plus une plante"herbivore". La grande surface collante de ses feuilles permet à cette espèce de capter des grains de pollen en quantité. Ils sont ensuite digérés, cela d'autant plus facilement que les grains peuvent germer sur le mucilage. En effet la paroi du tube pollinique est beaucoup moins résistante et imperméable aux enzymes que le revêtement cireux qui recouvre le grain de pollen.
La fécondation croisée favorise la quantité de graines produites et leur viabilité. Les insectes sont à la fois pollinisateurs et proies des plantes carnivores. En règle générale, le régime carnivore ne pertube pas la production de graines. La position des fleurs éloignée des pièges ou bien la date d'apparition des fleurs évite généralement un conflit entre nutrition et reproduction.
